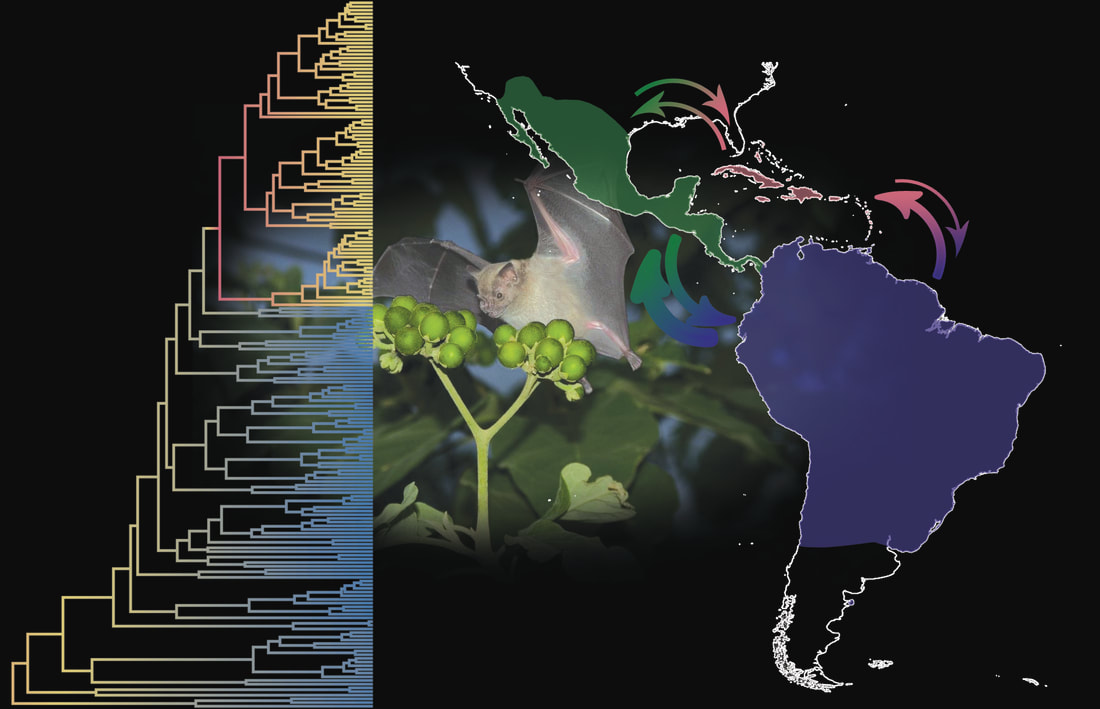
PHYLLOSTOMID BATS
Home
Book Table of Contents
About us
Home
Book Table of Contents
About us